Article Text

Abstract
The BCG vaccine protects non-specifically against other diseases than tuberculosis. Three randomised controlled trials of early BCG in Guinea-Bissau found a 38% reduction in all-cause neonatal mortality. Little is known about the underlying mechanisms. In Guinea-Bissau, prevalent infectious diseases display distinct seasonality. Revisiting the three trials (>6500 infants) comparing early BCG versus no early BCG in low weight infants on all-cause neonatal mortality over 12 consecutive years, we explored the seasonal variation in BCG’s effect on mortality. In a subgroup of participants, adaptive and innate cytokine responses were measured 4 weeks after randomisation. Consistently over the course of the three trials and 12 years, the effect of BCG on all-cause neonatal mortality was particularly beneficial when administered in November to January, coincident with peaking malaria infections. During these months, BCG was also associated with stronger proinflammatory responses to heterologous challenge. Recent studies have suggested a protective effect of BCG against malaria. BCG may also ameliorate immune-compromising fatal effects of placental malaria in the newborn.
- immunisation
- maternal health
- paediatrics
- vaccines
- malaria
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key questions
What is already known?
BCG vaccine non-specifically protects against other diseases than tuberculosis presumably via its immune-modulatory properties, training the innate immune system to enhance response potential.
Malaria in pregnancy is associated with an immune-compromised phenotype in the newborn and a higher infant morbidity and mortality.
What are the new findings?
Revisiting three randomised controlled trials of early BCG vaccination in low weight babies in Guinea-Bissau, we found that consistently over the course of the three trials spanning 12 years, the protective effect of BCG on neonatal mortality was particularly observed when administered in November to January, with modest effects in the remaining months.
This seasonality was coincident with the monthly malaria incidence pattern.
The immunological effects of BCG on proinflammatory adaptive and innate in vitro cytokine responses were stronger in the same months.
What do the new findings imply?
We hypothesise that neonatal BCG vaccination ameliorates the detrimental immunological effects of malaria in pregnancy in the newborn, thereby increasing resilience against infectious insults in early life.
Introduction
Besides protecting against disseminated tuberculosis (TB), the BCG vaccine has beneficial non-specific effects (NSEs), reducing mortality to other infections.1 2 In Guinea-Bissau, it was customary not to provide BCG to low weight or preterm infants. We therefore had the opportunity to investigate the effect of providing early BCG to low weight infants in three randomised controlled trials (RCTs) from 2002 to 2013. The intervention group received BCG at the first possible vaccination contact after birth; the control group received BCG according to local practice, usually around 6 weeks of age.2–4 The trials included a total of more than 6500 newborns.
All three trials found that early BCG reduced all-cause mortality; the combined meta-estimate showed a 16% reduction (mortality rate ratio (MRR) 0.84 (0.71–1.00)) in infant mortality (12 months) by early BCG and a very marked 38% reduction in neonatal mortality (28 days) (MRR 0.62 (0.46–0.83)).2 Verbal autopsy data and hospital records suggest that BCG particularly reduced neonatal mortality due to septicaemia and respiratory infections.2 5
Recent immunological studies have suggested that the beneficial NSEs of BCG may be linked to epigenetic changes of the innate immune system.6
Tropical Guinea-Bissau in West Africa has two distinct seasons defined by precipitation patterns. Although there is some variation between different years, we have usually classified the rainy season from June to November and the dry season from December to May. A number of important infectious diseases endemic to children in Guinea-Bissau and neighbouring countries follow a seasonal pattern, including rotavirus,7 respiratory syncytial virus (RSV),8 9 cryptosporidium10 and malaria.11 We speculated that if BCG non-specifically affects susceptibility to infections with a seasonal prevalence pattern, the BCG effect on neonatal mortality might be dependent on season.
In an explorative analysis, we therefore investigated potential seasonal variation of the beneficial NSEs of BCG on neonatal mortality.
Methods
Trial site and design
The Bandim Health Project (BHP) has conducted three RCTs of early BCG vaccination of low birth infants (<2.50 kg) in urban Bissau, Guinea-Bissau, in 2002–2004 (Trial I),3 2004–2008 (Trial II)4 and 2008–2013 (Trial III).2 In brief, recruitments took place at the maternity ward at the national hospital in capital Bissau, and three private hospitals in the capital, or for infants born at home when they came for their first vaccination at one of the three health centres in the BHP health and demographic surveillance site study area or at immunisation days at a private hospital. Infants weighing less than 2500 g at the time of assessment, without major malformations, who were not overtly sick, and had not received BCG, were eligible to participate in the trials.
Randomisation
Boys and girls were randomised separately (1:1) in blocks of 24 to receive early BCG, that is, at discharge or first consultation, or delayed BCG. Infants allocated to the early BCG group were vaccinated intradermally in the upper deltoid region by trained nurses with 0.05 mL BCG vaccine SSI (also known as BCG-Denmark, strain 1331, Statens Serum Institut (SSI), Copenhagen, Denmark). The children who were allocated to delayed BCG were treated according to local practice, that is, their mothers were told to have their children BCG vaccinated at the local health centres when the child had obtained a weight >2500 g; in practical terms, the mothers typically waited until 6 weeks of age when the child was due to receive the first diphtheria-tetanus-pertussis-containing vaccine and second oral polio vaccine (OPV). All children received OPV at birth.
Follow-up
After enrolment, the children and their mothers and/or guardians were driven home.
Infants were visited at home by the field team 3 days and 2 months after enrolment in order to assess neonatal mortality, and again at 6 and 12 months of age. When a death was registered, the assistant asked for the child’s vaccination card. A verbal autopsy was conducted about 3 months after the death by a trained assistant. A local doctor reviewed the autopsy and assigned a diagnosis.
Immunological study
Within Trial III, 467 infants randomised to early BCG or delayed BCG from March 2011 to December 2011 were included in an immunological subgroup study and had in vitro cytokine responses to innate agonists and recall antigens assessed 4 weeks after randomisation to assess the effect of early BCG on in vitro cytokine responses to purified protein derivative (PPD) and innate agonists.12 In brief, for enrolment into the immunological subgroup study, infants were visited at home by the study team 4 weeks after randomisation to early BCG or delayed BCG. Provided informed consent, blood was collected by heel puncture into a heparinised tube. The blood was diluted 1:9 with supplemented RPMI-1640 and stimulated for 24 hours in a 37°C humidified incubator with 5% CO2 with the following (final concentration in parenthesis): phorbol 12-myristate 13-acetate (PMA) (100 ng/mL; Sigma-Aldrich) and ionomycin (1 µg/mL) (Sigma-Aldrich) as a positive control; PPD from Mycobacterium tuberculosis (Statens Serum Institut, Copenhagen, Denmark) (10 µg/mL) to assess the mycobacterial specific response; lipopolysaccharide (LPS) (10 ng/mL) (Sigma-Aldrich) (a Toll-like receptor (TLR) four agonist); (S)-(2,3-bis(palmitoyloxy)-(2-RS)-propyl)-N-palmitoyl-(R)-Cys-(S)-Ser-(S)-Lys4-OH,trihydrochloride (Pam3CSK4) (1 µg/mL) (a TLR2/1 agonist) (InvivoGen); thiazoloquinoline compound (CL075) (1 µg/mL) (a TLR8/7 agonist) (InvivoGen). Supernatants were kept at −70°C until analysis at SSI, Copenhagen, by an immunobead-based multiplexed assay as previously described.13 The cytokines analysed were interleukin (IL)-1β, IL-5, IL-6, IL-10, IL-17, interferon (IFN)-γ and tumour necrosis factor (TNF)-α.
Patient and public involvement
Participating mothers were not individually involved in the design or conduct of the study during the course of the study. The results of the three clinical trials have been reported to the Ministry of Health in Guinea-Bissau, who has since adopted BCG vaccination at birth to low birth weight infants as official national policy.
Statistical analysis
Neonatal mortality rates were compared for infants randomised to early or delayed BCG in a Cox proportional hazard model, with age as the underlying time. We adjusted the SE for the dependence of twins by using robust SE. In the analysis combining trials, we allowed separate baseline hazards for each trial. Season or month of enrolment was defined by the date of randomisation.
Cytokine data were log transformed. For several of the cytokines, a proportion of the measurements was below the lower limit of detection or above the upper limit of detection of the assay, and therefore denoted as non-detectable (ND) measurements. ND measurements were handled by multiple imputations based on Tobit regression.14 15 The parameters of the underlying normal distribution are estimated by Tobit regression and values below or above the detection limit are drawn multiple times from a truncated normal distribution. Geometric mean ratios (GMRs) were obtained as anti-logged coefficients. For distributions with >50% ND measurements, the proportion of detectable samples was analysed by Poisson regression giving proportion ratios (PRs).
Statistical analyses were performed using Stata V.12 software.
Results
Combining the three RCTs, in total 6525 infants and 189 neonatal deaths were included in the neonatal mortality analysis as previously published.2 Cohort characteristics have previously been described; background parameters were evenly distributed between the two randomisation arms2–4; age at randomisation and distribution of sex did not vary by month of enrolment in the two study arms; and distribution of age of reception of BCG in the delayed group did not vary by month (data not shown).
The overall neonatal mortality rates declined from Trial I over Trial II to Trial III.2 The number of deaths in Trial I was too small to be analysed separately by calendar month (online supplementary table 1). The monthly fluctuations in the crude neonatal mortality rates (by month of randomisation) were larger in Trial II than Trial III, which may be partly explained by the latter trial comprising a larger number of individuals (normalisation effect). Nonetheless, in both Trial II and Trial III, there were marked drops in the delayed BCG group mortality in July and October, and surges in August and December (online supplementary figure 1), whereas for the early BCG group in both trials, mortality rates were relatively high in July, and low in November, December, January and April (online supplementary figure 2).
Supplemental material
Comparing the neonatal mortality rates in the early BCG group with the delayed BCG group (online supplementary figure 3), we estimated HR by calendar month of randomisation for Trial II, Trial III and the three trials combined (online supplementary table 1; figure 1). Across all three trials and 12 years, there was a consistent pattern of a protective effect of BCG particularly during November to January. In contrast, there seemed to be an increased relative hazard after BCG in July. The effect estimate of early BCG seemed to covary inversely with the proportion of low birthweight children born in the BHP study area (the main catchment area and recruitment sites of the trials), with a nadir in July and more deliveries in October to March (online supplementary figure 4).

HR for neonatal death comparing early BCG with delayed BCG, per calendar month and by trial. For Trial II, no HR estimate for January and July due to no deaths in the early BCG group in January and delayed group in July, respectively. HR<1 indicates lower mortality in the BCG group; HR>1 indicates lower mortality in the delayed BCG group.
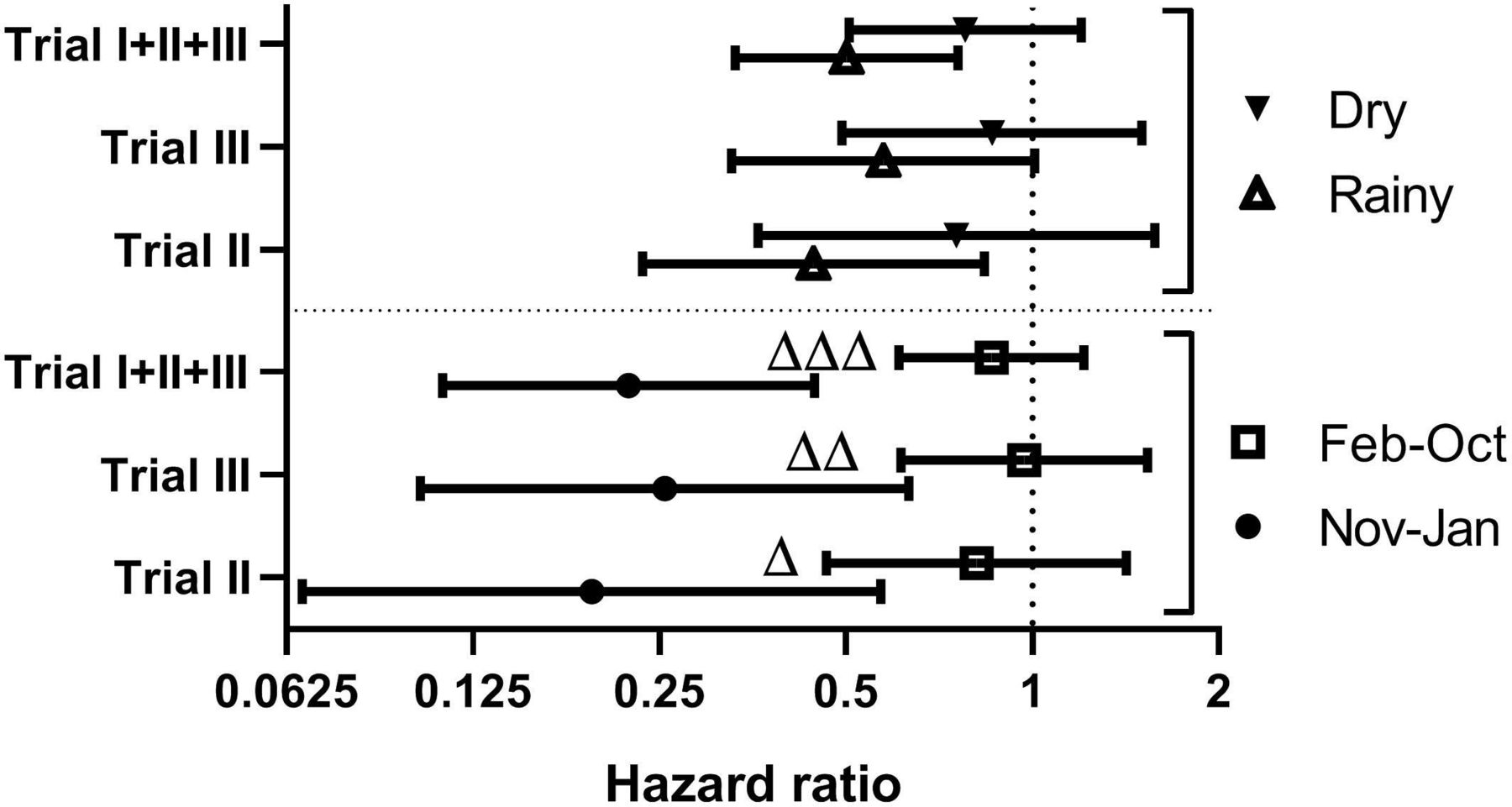
The monthly combined effect estimates (online supplementary table 1) differed significantly (test of heterogeneity, p=0.05). Applying the conventional seasonal dichotomy defined by precipitation (rainy season June to November; dry season December to May), the BCG effect estimates were not significantly different (test of heterogeneity, p=0.15) between the dry season (HR=0.50 (0.33–0.76)) and the rainy season (HR=0.78 (0.51–1.20)) (figure 2; online supplementary table 2).

HR for neonatal death comparing early BCG with delayed BCG, stratified by sex, trial and season. Estimated HR (with 95% CI) comparing early BCG to delayed BCG, stratified by season and trial. Season of BCG administration is defined as November to January versus February to October or dry (December to May) versus rainy (June to November). A HR of 1 is indicated with a vertical hatched line. Δp<0.05; ΔΔp<0.01; ΔΔΔp<0.001 for interaction between season and BCG.
In public health, the seasonal definition has often been a proxy for a high burden of malaria infection. In Guinea-Bissau, malaria is now a late rain-early dry season epidemic.11 Data from Bissau show that the malaria transmission in <15-year-old children mainly takes place from September to January (online supplementary figure 5). Comparing BCG effects in the period with high malaria transmission (September–January) with low malaria transmission (February–August) yielded significant effect modification, with a HR=0.41 (0.25–0.66) for September–January and a HR=0.83 (0.56–1.23) for February–August, p=0.02 for interaction between season and BCG (online supplementary table 3).
Letting the data define the annual stratification as November to January versus February to October, we found highly significantly different estimates of the effect of BCG on all-cause neonatal mortality (p=0.001), with a HR=0.22 (0.11–0.45) for November to January and HR=0.86 (0.61–1.21) for February to October (table 1; figure 2). This pattern was comparable in boys and girls. The stronger beneficial effect of BCG in November–January versus February–October was seen across all the study years (online supplementary figure 6).
Neonatal mortality rates and HR comparing early BCG with delayed BCG, by sex, trial and season (November–January vs February–October)
Triangulation
The observed pattern could indicate that BCG ameliorated the negative effects of maternal malaria infection late in pregnancy. To test this hypothesis, we performed several analyses to test the derived deductions.
Deduction 1: If maternal malaria late in pregnancy is harmful to the child, then there should be a negative association between being born by a mother exposed to high malaria transmission months and neonatal mortality in the control group, which should be abrogated in the BCG group. Indeed, malaria incidences in the month prior to randomisation was associated with a significant increasing risk of neonatal death in the delayed BCG group (HR=1.11 (1.00–1.22)), but a trend for an opposite effect in the early BCG group, resulting in an interaction between malaria incidences and BCG (p=0.03) (online supplementary table 3).
Deduction 2: If maternal malaria late in pregnancy is harmful to the child and primigravidae are more susceptible to malaria in pregnancy, then the protective effect of BCG should be strongest in primigravidae. We could confirm this deduction, as the seasonal differences in the effect of BCG was larger among newborns from primigravidae pregnancies (November–January: HR=0.05 (0.01–0.34) and February–October: HR=1.15 (0.68–1.94), p=0.002 for an interaction between season and BCG for the primigravidae), whereas the BCG effect estimates were not different between seasons for the multigravidae (p=0.26) (online supplementary table 3).
Cytokine responses
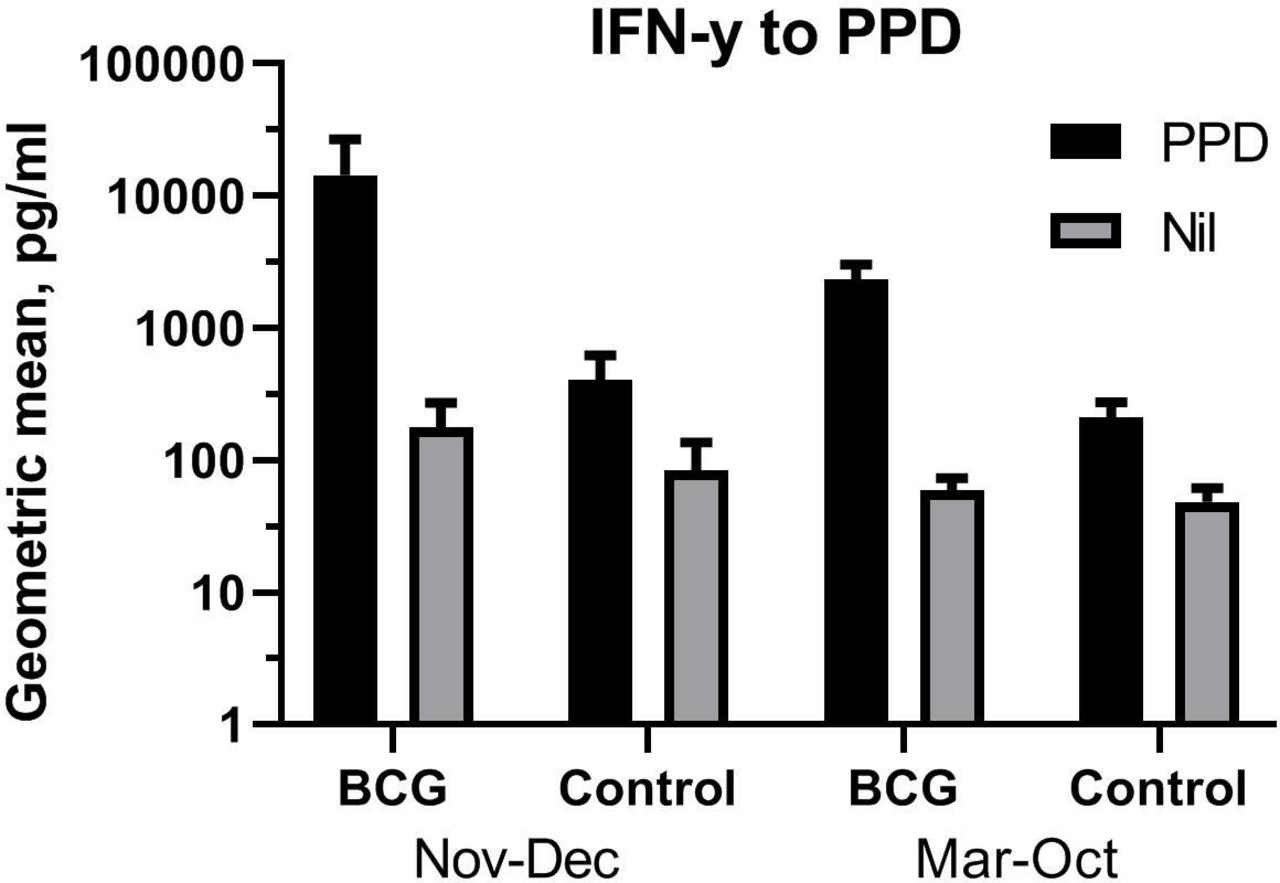
Within Trial III we had found that BCG overall increased innate cytokine responses of particularly IL-1β, IL-6, TNF-α and IFN-γ.12 Reanalysing the cytokine responses according to the seasonal dichotomy defined by the mortality patterns, comparing early BCG with control infants, the enhancing effect of BCG was stronger when administered in November to January compared with March to October (online supplementary table 4). This was seen for both specific responses of IFN-γ response to PPD stimulation (figure 3) and non-specific responses (online supplementary table 4). In a collective analysis combining the non-specific cytokine responses in a single estimate, BCG was associated with significantly larger IL-1β, IL-6, TNF-α, IFN-γ and IL-17 responses to heterologous challenge in November to December compared with the other months (figure 4).

Interferon (IFN)-γ responses to purified protein derivative (PPD) and medium alone. Geometric means of IFN-γ concentrations in whole blood in vitro stimulated with PPD (10 μg/mL) 4 weeks after randomisation to early BCG or nothing (control), stratified by season (November to December vs March to October). Error bar is 95% CI.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
The immunological effects of BCG on in vitro cytokine responses (collective test). Whole blood from infants randomised to early BCG or nothing was collected 4 weeks after randomisation and stimulated with a panel of innate agonists and recall antigens (Jensen et al, J InfectDis 2015 211: 956–967), stratified by time (calendar month) of enrolment. The geometric mean ratios (GMRs) comparing early BCG vaccinated with non-vaccinated infants were analysed collectively for the non-specific responses to LPS, Pam3CSK4, PMA/ionomycin, CL075 and medium alone using Tobit regression. The study included infants randomised from March to December 2011. Season defined by the month of randomisation as November–December, n=101 versus March–October, n=366. *p<0.05; ***p<0.001 for test of BCG; ΔΔp<0.01 for interaction between season and BCG.
Discussion
Reanalysing three RCTs with similar design covering a period of 12 years, we found that the protective NSE of early BCG vaccination on all-cause neonatal mortality in a tropical low-income setting was season-dependent, showing a consistent pattern of the most pronounced beneficial effects in November to January, corresponding to the late rainy and early dry season, coinciding with the months when most expecting mothers would be exposed to malaria. This effect was seen across all three trials and across all study years.
Strengths and weaknesses
The present analysis was not preplanned. As the immunological study was conducted over less than a year, spanning from April 2011 to February 2012, it does not in its own right support conclusions on seasonal effects. Nevertheless, it is noteworthy that the findings on the effect of BCG on mortality are reflected in the immunological effects on cytokine responses in the subcohort.
Seasonal variation in mortality
A number of child health studies in Guinea-Bissau have investigated mortality risk by season, applying the conventional seasonal definition based on rain patterns. Accumulated data from repeated population surveys in rural Guinea-Bissau covering the period from 1990 to 2013 show that while the overall mortality steadily declined, the under-five child mortality remained considerably higher in the rainy season than in the dry season throughout the study period, averaging 51% higher (95% CI 45% to 58%).16
In urban Bissau, the seasonal difference was also evident although less pronounced in 1991 to 1996 with 15% (95% CI 4% to 28%) higher under-five mortality in the rainy season versus the dry season.17 Also, hospitalisation rates and in-hospital deaths were higher in the rainy season.
Seasonal variation in pathogen exposure
A number of important pathogens infecting children display seasonal variation. In a community-based study in Bissau, all-cause diarrhoea in <4-year-old children was considerably higher in the rainy season. The parasite Cryptosporidium parvum is one organism causing diarrhoea. In Guinea-Bissau, it was most prevalent in the early rainy season and an important cause of death in children less than 1 year.10 Another diarrhoea-causing enteropathogen, rotavirus, with importance to young infants,18 also exhibits a seasonal pattern, with annual epidemics occurring during the relatively dry and cooler months, from January to April.7
Studies of the aetiology of respiratory infections in young children from Senegal19 and Cameroon20 identify adenovirus and RSV as the most common pathogens in infants, whereas a case–control study of severe acute respiratory infections in Kenyan children found that influenza and RSV were the largest risk factors for being a respiratory infectious case.21 Recently, a large multicentre survey in under-five hospital admissions for severe pneumonia in seven African and Asian countries found that RSV was the leading cause of respiratory hospitalisation across all study sites, partaking 31% of the infectious fraction.22
A previous study found that BCG vaccination was associated with reduced RSV hospitalisation in children.23 Although there is a paucity in data on RSV from Guinea-Bissau, a study from neighbouring The Gambia found that RSV diagnosed hospitalised cases peaked in August to October,8 whereas in Senegal, the RSV season in children was mostly in July to November.9
In conclusion, the seasonal variation observed in the above pathogens does not seem to explain the seasonal variation in the BGC effect; most of the pathogens known to fluctuate with season would not cause neonatal mortality and the RSV epidemic seems to peak before the period where we see the strongest beneficial BCG effect.
In Guinea-Bissau, malaria has gradually shifted from being endemic to epidemic particularly in October to December. BCG has been reported to protect against malaria. A murine study found that BCG 2 months prior to challenge with Plasmodium yoelii lowered the ensuing parasitaemia.24 In Guinea-Bissau, Roth et al found a reduction in malaria mortality for children with a BCG scar.25 Recently an observational study in under-five children using data from several sub-Saharan African countries found that BCG-vaccinated children had a lower risk of having a positive rapid diagnostic test (Berendsen et al, submitted).
In a recent human experiment, using a controlled human malaria infection model, malaria parasitaemia and clinical symptoms were compared in BCG-vaccinated and non-vaccinated adults. In half of the BCG-vaccinated volunteers, malaria parasitaemia was reduced, concomitant with more pronounced clinical symptoms, increased early innate responses to the malaria challenge, increased plasma inflammatory IFN-γ, granzyme B and C reactive protein, and increased cytolytic activity of natural killer (NK) cells in response to Plasmodium falciparum, compared with non-vaccinated control subjects.26 The study hence indicated that BCG vaccination via mechanisms reminiscent of trained immunity6 could alter the clinical and immunological response to malaria infection. Of note, most of the traits of trained immunity associated with BCG were only observed after the malaria challenge, not in the vaccinated but homeostatic phase prior to malaria.
Malaria in pregnancy is a risk factor of congenital malaria, the result of transplacental transmission of parasites. Epidemiological studies in sub-Saharan Africa have produced variable estimates of the rate of congenital malaria.27 An observational study from Nigeria found that of 546 newborns admitted to a local neonatal intensive care unit for suspicion of sepsis, 13% had congenital malaria.28 However, in a large cohort study among all neonates living in a malaria endemic region and admitted to a district hospital in Kenya during 2002–2009, only 1% had P. falciparum malaria parasitaemia; the case fatality rate of those with parasitaemia was slightly but non-significantly higher than non-parasitemic newborns.29 In the immunological study during 2011–2012, no malaria was found by microscopic examination in infants 4 weeks after randomisation,12 indicating that neonatal malaria was rare.
Malaria in pregnancy, irrespective of congenital malaria, is a risk factor for low birth weight, neonatal mortality30 and reduced infant growth,31 and placental malaria has been associated with an increased susceptibility to malaria during infancy.32 33 Moreover, a community study in Benin with a prevalence of placental malaria of 11% found an association with increased incidences of non-malaria febrile episodes (including both gastrointestinal and respiratory symptoms) during the first 18 months of life (adjusted incidence rate ratio 1.4 (1.1–1.8),34 indicating a generalised long-lasting immune-compromising effect of placental malaria in the infant, even though the infant may not be infected.
Interpretation of the epidemiological evidence
Triangulating on the evidence of BCG vaccination providing health benefit to infants exposed to malaria in pregnancy, the following can be deducted from the present data and circumstantial evidence: (1) We found a strong variation in BCG effect on neonatal mortality by calendar months, with most beneficial effects during November–January, months during which mothers were most likely to be exposed to malaria. (2) Whereas the monthly estimated malaria incidences in <15-year-old children in Bissau during the study period ranged from 0 to 8.3 per 1000 persons/month,11 the epidemiology of pregnancy malaria in our BCG trial cohorts was unknown. A survey using microscopic examination of peripheral blood from pregnant women in the study area in 2001–2002, leading up to the initiation of Trial I, found that the malaria parasitaemia prevalence was 57%.35 Hence, although the survey was not contemporary to our BCG trial, and the malaria epidemic has changed over the course of the BCG trials,11 the findings indicate relevance of malaria in pregnancy to the present cohorts. A study in the neighbouring country The Gambia in 2010–2011 among pregnant women attending antenatal care found a malaria prevalence of 9%.36 (3) Malaria in pregnancy is a risk factor for low birth weight,37 which increases the probability of the exposure in our low birthweight cohort. (4) Compiling data from four African sites, the prevalence of pregnancy malaria increased with the number of months of the pregnancy taking place in the high malaria transmission season.36 Malaria in pregnancy is a risk factor for neonatal death.37 We found indication that malaria transmission rates in the month prior to enrolment was associated with increased neonatal mortality in the delayed BCG group, whereas this was not seen in the early BCG group. (5) Primigravidae women experience a higher risk of pregnancy malaria than multigravidae, and the association between placental malaria and low birth weight or neonatal death is stronger in primigravidae (reviewed in previous work37). Indeed, we found that the seasonal effect modification was particularly evident in infants born to primigravidae women.
The beneficial NSEs of BCG are of variable magnitude.4 38 A number of potential explanations are in play, including differences in BCG strain,39–41 batch variation strains,42 the enhancing effect of parental priming with BCG,43 44 genetic polymorphisms45 46 and different underlying aetiologies of neonatal disease and death. The epidemiological data presented here suggest that season could be a potential effect modifier, probably driven by underlying temporal dynamics of the pathogen burden.
Studies from some low-income countries have suggested that changes in nutritional status could contribute to the seasonality in morbidity and mortality47 or differential vaccine responses.48 However, in the present setting urban districts of the capital Bissau, there is limited seasonal difference in overall food availability. While we cannot exclude that there could be interaction between BCG and certain micronutrients, which may differ in availability by season, we find it unlikely that such interaction would explain the observed seasonal variation of BCG effects.
Immunological effects on the newborn of malaria exposure in pregnancy
Immunological studies have indicated that malaria in pregnancy alters the immunological phenotype in the newborn. Particularly, decreased inflammatory responses and increased frequencies of regulatory T cells have been observed. Infants born with placental malaria had a lower frequency of IFN-γ producing CD4+ T cell in response to PPD 12 months after BCG at birth compared with infants with no placental malaria.49
Cord blood from Gabonese babies born with placental malaria (parasitemic placental blood) produced more IL-10 in response to non-specific CD3/CD28 polyclonal stimulation compared with placental malaria-negative newborns,50 although the IFN-γ response of cord blood to phytohaemagglutinin (PHA) stimulation did not differ significantly by malaria exposure.51 In The Gambia, cord blood mononuclear cells from babies born with parasitemic placentas produced less IFN-γ and more IL-10 to PHA stimulation,52 less IL-12p40 to LPS53 compared with cord blood from births with non-parasitemic placentas. Moreover, acute placental malaria or histological evidence of past placental malaria was associated with an increased frequency of IL-10+CD4+ T cells in cord blood in response to Staphylococcal enterotoxin B (SEB).54 Also, specific responses to P. falciparum were reduced for IFN-γ and increased for IL-10, respectively, in placental malaria-positive infants’ cord blood.52 Depletion of CD4+CD25+FoxP3+ cells52 or addition of LPS53 partly restored the reduced IFN-γ responses to PHA, as well the reduced specific responses to P. falciparum.51 52 Furthermore, in the cord blood from the above Gabonese study, a subset of CD4+CD25+ T cells exerted repressive effects on monocyte MHC class I and II expression.50
Kenyan infants born to mothers malaria positive at delivery could be segregated into responders versus non-responders to malaria blood stage antigens by IFN-γ, IL-2, IL-13 and/or IL-5 production by cord blood mononuclear cells. Non-responders nevertheless had an increased IL-10 response to malarial antigen stimulation (hence evidence for prenatal malaria immune experience) compared with both the responding infants and (control) infants born to malaria-negative mothers. The malaria antigen-specific hyporesponsiveness could be partly reversed by co-culturing with the clonal stimulating cytokines IL-2 +IL-15, indicative of the exposed non-responders had adopted an immune tolerant phenotype. The phenotype of the malaria exposed but non-responding infants characterised by a lower frequency of malaria antigen-driven IFN-γ and/or IL-2 production and higher IL-10 release persisted at 6-month follow-ups, and in turn, these infants also had an increased risk of malaria infection compared with unexposed controls and exposed responders.55
Cord blood from Cameroonian infants born with placental P. falciparum malaria had a reduced frequency of Vδ2 (γδ) lymphocytes, a shift towards a central memory phenotype of Vγ2Vδ2 lymphocytes and a reduced proliferative capacity of certain common γδ T cell clones compared with unexposed infants, indicative of placental malaria selectively exhausting the γδ repertoire.56 Noteworthy, BCG vaccination of human newborns was found to expand the γδ T cell population,57 and the γδ T cells together with NK cells are the most vivid producers of anti-mycobacterial IFN-γ in vitro in BCG-vaccinated infants.58
A few studies have investigated whether the immune-modulating effect of malaria in pregnancy depends on the timing of exposure during pregnancy and whether the placental malaria (as determined by histological examination of the placenta) is resolved (past), acute (ongoing) or chronic. The evidence is somewhat conflicting.
In Gambian cord blood, chronic and resolved placental malaria cases but not for acute malaria (placental parasitaemia at delivery) had increased IFN-γ, TNF-α secretion and IFN-γ:IL-10 ratio after SEB stimulation, as well as an increased frequency of regulatory T cells (CD4+CD25+FoxP3+) compared with placental malaria-negative cases.54
Likewise, in a study of Ugandan mother–infant pairs, cord blood FoxP3 +T regulatory cell counts were higher in infants born to mothers with Plasmodium parasitaemia early in pregnancy, but not in infants with placental parasitaemia at the time of delivery.59 The effect was particularly seen in males.60 In contrast, there was a higher frequency of activated CD4+ T cells (CD25+FoxP3-CD127+) in the cord blood of neonates with active placental Plasmodium infection at the time of delivery but not earlier during the pregnancy.59 Compared with infants born to mothers with at least one episode of treated malaria during the pregnancy, but non-parasitemic at delivery, cord blood from infants born with parasitemic placentas produced less IL-2 in response to CD3/CD28 stimulation.50 In Burkina Faso, evidence of malaria in pregnancy, irrespective of whether the malaria exposure was acute, chronic or resolved, was associated with decreased innate spontaneous (non-stimulated) proinflammatory as well as anti-inflammatory cytokine and chemokine production in cord blood.61
Interestingly, in Tanzanian newborns, increased TLR3-mediated and TLR7/8-mediated anti-inflammatory IL-10 responses in cord blood were associated with a significantly higher risk of subsequent P. falciparum infection in infancy.62
In summary, the above studies indicate that malaria exposure in pregnancy may drive a more tolerant and less responsive immune phenotype in the newborn, which may persist throughout infancy and pose the infant of infectious risk. To that end, BCG has potent immune-stimulatory properties, including adjuvant-like functions potentially enhancing immune responses to other concomitant vaccinations in adults as well as infants (reviewed in previous work63); BCG is capable of inducing Th1 directed responses in the newborn,64 including polyfunctional vaccine-specific T cells responses.65 Studies of the general non-specific immunological profile induced by BCG in newborns find indication of increased innate responses after BCG in non-stimulated whole blood, including the present cohort from Guinea-Bissau,12 66 whereas analyses of the responses to innate stimulation has produced more mixed results, indicating reduced responses of IL-6, macrophage inflammatory protein (MIP)-1α and MIP-1β to TLR2 and TLR7/8 stimulation, but also reduced anti-inflammatory responses of IL-10 and IL-1 receptor agonist in Australian newborns 1 week after BCG,66 whereas there was no or negligible effects of BCG in Danish newborn babies evaluated at 4 days and 3 months after BCG.67 In the present cohort from Guinea-Bissau, BCG was associated with increased proinflammatory responses in PMA-ionomycin and TLR2 stimulated whole blood and reduced IL-10 responses to TLR7 agonists at 1 month of age.12 On a functional level, observational studies from Guinea-Bissau using skin prick tests in infants and young children have indicated a protective effect of BCG vaccination against anergy (non-responsiveness) to common (heterologous) vaccine antigens.68 Interestingly, anergy in infants seemed more prevalent in the rainy season,68 69 concomitant with supposedly increased malaria exposure. It could therefore be speculated if BCG also after the neonatal period (perhaps administered as a booster vaccine) may reverse malaria-induced anergy and associated mortality in the rainy season.
Conclusion
The enhanced proinflammatory effects of BCG on cytokine levels in Guinean infants born during the peaking malaria season, the enhanced beneficial effect on mortality in the same season, and the well-known reduced inflammatory potential as well as increased regulatory immune phenotype of babies exposed to malaria in pregnancy converge to a coherent hypothesis. We propose that BCG may ameliorate the detrimental effect of malaria exposure, thereby strengthening the ability of the newborn to respond to infectious challenges in early life. These observations should be tested for reproducibility in other relevant settings.
Acknowledgments
We thank the mothers and infants who participated in the study.
References
Footnotes
Handling editor Seye Abimbola
Contributors KJJ, SB-S, PA and CSB conceived and designed the BCG-related studies. JU and P-ELK carried out the malaria surveys. KJJ and SB-S analysed the data. KJJ wrote the first draft of the manuscript. All authors contributed to the final version of the manuscript.
Funding The randomised controlled trials of early BCG were supported by The European Research Council (starting grant ERC-2009-StG-243149); the Novo Nordisk Foundation (research professorship grant to PA); the Danish National Research Foundation (grant DNRF108 to the Research CenterCentre for Vitamins & Vaccines); and DANIDA, European Union FP7, and OPTIMUNISE (grant Health-F3-2011-261 375 to the Bandim Health Project). The immunological subgroup study was supported by Novo Nordisk Foundation and University of Southern Denmark; KJJ is supported by a grant from Novo Nordisk Foundation (grant NNF14OC0012169).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The BCG RCTs and the immunological substudy were approved by the National Committee on Health Ethics of the Ministry of Health in Guinea-Bissau; a consultative approval was obtained from the Danish National Committee on Biomedical Research Ethics. The BCG trials were registered with clinicaltrials.gov, numbers NCT00146302 (Trials I+II), NCT00625482 (Trial III).
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available on reasonable request.